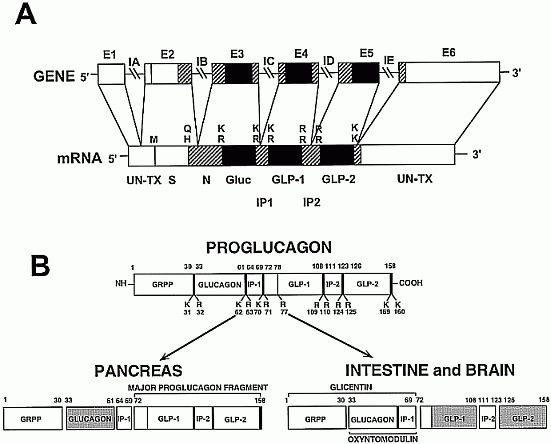
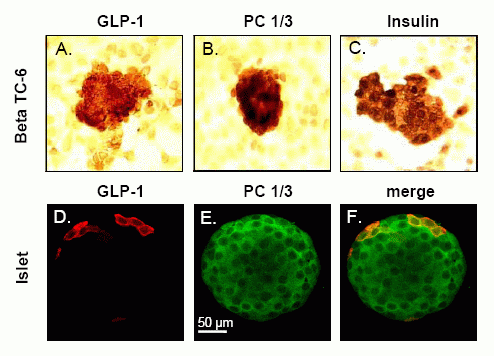
Expression of the preproglucagon gene. A, Diagram of the
proglucagon gene and encoded mRNA. B, Alternative posttranslational processing
of proglucagon in pancreas, intestine, and brain.
Structure of proglucagon and biological actions of GLP-1 and GLP-2.
The principal target cell types for GLP-1 (islet ß cells stained with insulin
antiserum) and GLP-2 (intestinal endocrine cells stained with GLP-2 receptor antiserum) are shown below the peptide sequences. The biological actions of
the peptides are summarized below. GRPP, Glicentin-related pancreatic polypeptide; IP, intervening peptide; GLP-2R, glucagon-like peptide-2
receptor.
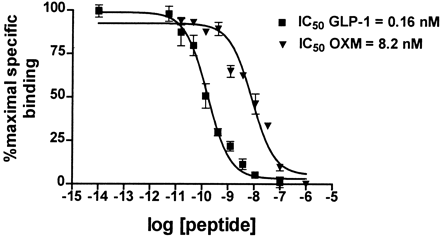
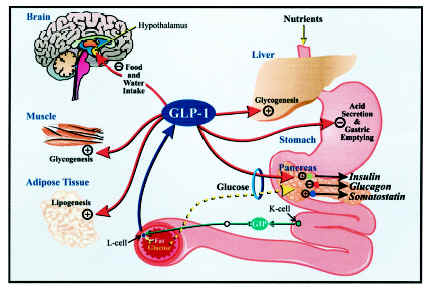
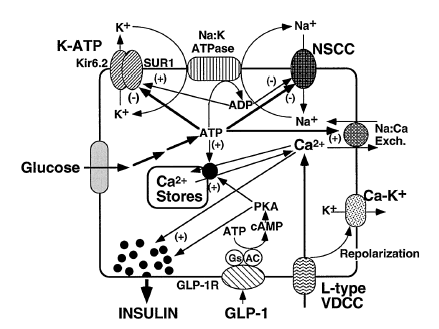
Glucagon-like peptide-1 (GLP-1) is derived from the transcription product of the proglucagon gene. The major source of GLP-1 in the body is the intestinal L cell that secretes GLP-1 as a gut hormone. The biologically active forms of GLP-1 are: GLP-1-(7-37) and GLP-1-(7-36)NH2. Those peptides result from selective cleavage of the proglucagon molecule.
Mammalian Preproglucagon and the PGDPs
Structure of Mammalian Proglucagon and Tissue-Specific Processing of Proglucagon into the PGDPs. The numbers above and below the proglucagon sequence refer to specific
amino acid residues within proglucagon. IP, Intervening peptide.
Structure of mammalian proglucagon, proglucagon-derived peptides, and biological actions of GLP-2.
Structure of mammalian proglucagon, the proglucagon-derived peptides, and the biological actions of GLP-2. The numbers above and below the proglucagon molecule correspond to
the positions and length of specific PGDPs. MPGF, Major proglucagon fragment; IP, intervening peptide.
Central pre-proglucagon derived peptides: opportunities for treatment of obesity
Modern societies have moved from famine to feast and obesity and its co-morbidities now sweep the world as a global epidemic. Numerous scientific laboratories and pharmaceutical
companies have taken the challenge and are now exploiting novel molecular targets for treatment of obesity. The pre-proglucagon system constitutes interesting candidates as potential targets for
new anti-obesity drugs. In the periphery, pre-proglucagon derived peptides, Glucagon-Like Peptide-1 (GLP-1), Glucagon-Like Peptide-2 (GLP-2) and oxyntomodulin (OXM) are involved in a wide variety of
physiological functions, including glucose homeostasis, gastric emptying, intestinal growth, insulin secretion as well as the regulation of food intake. Peripheral administration of GLP-1
derivatives and analogues to both rodents and man have shown promising effects on food intake and body weight suggesting that such therapies constitute potential anti-obesity treatment. In the
central nervous system, pre-proglucagon and hence GLP-1, GLP-2 and OXM are exclusively found in a small population of nerve cells in the nucleus of the solitary tract. These constitute a neural pathway
linking the "viscero-sensory" brainstem to hypothalamic nuclei involved in energy homeostasis. Intracerebroventricular administration of all of the three derived peptides robustly
decrease food intake. It is evident that central GLP-1 agonism probably in combination with GLP-2 and/or OXM agonism constitute a potential pharmacological tool to reduce food intake and maybe also
enhance energy expenditure. This and other aspects of the current state of the role of central pre-proglucagon in energy homeostasis
are reviewed.
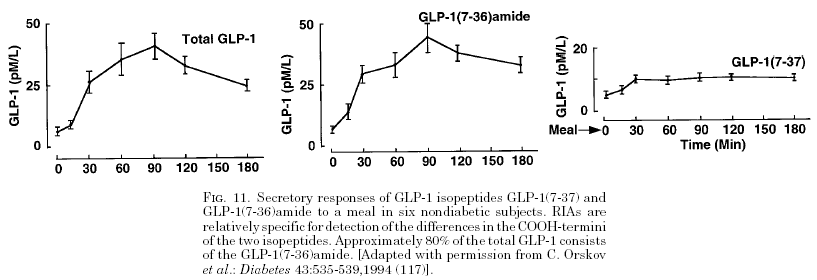
GLP-1 (9–36) Amide, Cleavage Product of GLP-1 (7–36) Amide, Is a Glucoregulatory Peptide
Objective:
Glucagon-like peptide-1 (GLP-1) (7–36) amide is a glucoregulatory hormone with insulinotropic and insulinomimetic actions. We determined whether the insulinomimetic effects of GLP-1 are mediated through its principal metabolite, GLP-1 (9–36) amide (GLP-1m).
Methods and Procedures:
Glucose turnover during two, 2-h, euglycemic clamps was measured in 12 lean and 12 obese (BMI <25 or >30 kg/m2) male and female subject volunteers with normal oral glucose tolerance test. Saline or GLP-1m were infused from 0 to 60 min in each study. Additionally, seven lean and six obese subjects underwent a third clamp in which the GLP-1 receptor (GLP-1R) antagonist, exendin (9–39) amide was infused from -60 to 60 min with GLP-1m from 0 to 60 min.
Results:
No glucose infusion was required in lean subjects to sustain euglycemia (glucose clamp) during saline or GLP-1m infusions. However, in obese subjects glucose infusion was necessary during GLP-1m infusion alone in order to compensate for a marked (>50%) inhibition of hepatic glucose production (HGP). Plasma insulin levels remained constant in lean subjects but rose significantly in obese subjects after termination of the peptide infusions. During GLP-1R blockade, infusion of glucose was immediately required upon starting GLP-1m infusions in all subjects due to a more dramatic reduction in HGP, as well as a delayed and modest insulinotropic response.
Discussion:
We conclude that GLP-1m potently inhibits HGP and is a weak insulinotropic agent. These properties are particularly apparent and pronounced in obese but only become apparent in lean subjects during GLP-1 (7–36) receptor blockade. These previously unrecognized antidiabetogenic actions of GLP-1m may have therapeutic usefulness.
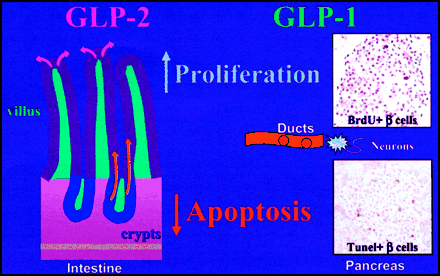
Proliferative and Antiapoptotic Effects of GLP-1 and GLP-2 in the Pancreas and Intestine, Respectively.
GLP-1 stimulates cell proliferation in pancreatic ductal cells and islets, and exerts antiapoptotic actions on islet ß-cells and neurons. GLP-2 stimulates intestinal crypt cell
proliferation and inhibits apoptosis in the crypt and enterocyte compartments of the gut epithelium.
Comparison of the effects of ICV and iPVN pro-glucagon-derived and related products
Comparison of the effects of ICV and iPVN proglucagonderived and related products on food intake in fasted rats. A, Cumulative food intake (grams) up to 8 h after
ICV injection of GLP-1, OXM, glucagon, or glicentin (all 3 nmol) into fasted animals. *, P < 0.05 vs. saline control, B, Cumulative food intake (grams) up to 24 h
after an acute iPVN injection of GLP-1, OXM (both 1 nmol), or exendin-4 (0.03 nmol) into fasted animals. *, P <
0.01 vs. saline control for all groups at 1, 2, and 4 h. *, P < 0.05 vs. saline control for exendin-4 only at 8 h.
Effects of ICV and iPVN OXM on food intake in fasted rats.
A, Cumulative food intake (grams) up to 8 h after an acute ICV injection of OXM (0.3, 1, 3, or 10 nmol). B, Cumulative food intake (grams) up to 8 h after an acute iPVN injection of OXM (0.1, 0.3, or 1.0 nmol) into fasted animals. *, P < 0.05 vs. saline control.
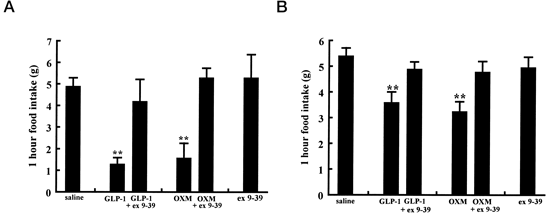
Inhibition of OXM and GLP-1 effects on food intake by exendin-(9–39).
A, Food intake 1 h after an acute ICV injection of GLP-1 (3 nmol), GLP-1 plus exendin-(9–39) (30 nmol), OXM (3 nmol), OXM and exendin- (9–39) (30 nmol), or exendin- (9–39) alone (30 nmol). B, Food intake after an acute iPVN injection of GLP-1 (1 nmol), GLP-1 and exendin-(9–39) (10
nmol), OXM (1 nmol), OXM and exendin-(9–39) (10 nmol), or exendin-(9–39) alone (10 nmol) into fasted animals. **, P < 0.005 vs. saline control.
Effect of ICV OXM at the onset of the dark phase. Sated rats received an ICV injection of OXM, GLP-1 (3 nmol), or saline at the onset of the dark phase. Food intake (grams; A) and behaviors (B) at 1 h postinjection were
determined. *, P < 0.05 vs. saline control.
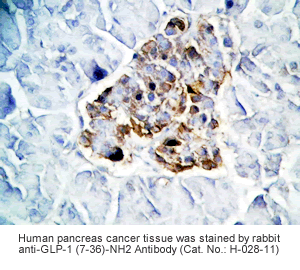
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min
Amplification
Streptavidin-HRP (Vector), 1:400, 30 min
Detection System
HRP
Substrate
DAB (Sigma), 3 min
Counterstained
Hematoxylin, 30 sec
Mapping in Rat Pancreas and Intestines by GLP-1 (7-37) Antibody (H-028-13)
Tissue Sample
Rat Pancreatic and intestinal tissue
Fixative
10% formalin
Embedding
Paraffin
Negative Control
Pre-immuno serum
Pretreatment
Intact
Blocking
2% Normal Goat Serum
Primary Antibody
Rabbit Anti-GLP-1 (7-37) (Human, Rat, Mouse, Porcine) Serum
(Catalog No.:H-028-13)
Optimal Dilution
1:500, 1 hour at RT
Secondary Antibody
Goat Anti-Rabbit IgG, Biotinylated (1:400), 30 min
Amplification
ABC (Vector) (1:400, 30 min)
Detection System
HRP
Substrate
DAB (Sigma), 3 min
Counterstained
with methyl green
Rat pancreatic and intestinal sections with an antiserum for human GLP-1 (7–37) (Phoenix Pharmaceuticals, Mountain View, California, USA) at a concentration of 1/500. The second antibody was a biotinylated goat anti-rabbit immunoglobulin, and the signal was detected using the Vectastain ABC kit (Vector Laboratories, Burlingame, California, USA) with DAB as a substrate. The section was counterstained with methyl green.
Granata et al. Obestatin promotes survival of pancreatic beta-cells and human islets and induces expression of genes involved in the regulation of beta-cell mass and function. Diabetes. 2008 Apr;57(4):967-79.
Santoro et al. Enterohormonal changes after digestive adaptation: five-year results of a surgical proposal to treat obesity and associated diseases. Obes Surg. 2008 Jan;18(1):17-26.
Pacheco et al. The effects of duodenal-jejunal exclusion on hormonal regulation of glucose metabolism in Goto-Kakizaki rats. Am J Surg. 2007 Aug;194(2):221-4.
Suzuki et al. Changes in GI hormones and their effect on gastric emptying and transit times after Roux-en-Y gastric bypass in rat model Surgery. 2005 Aug;138(2):283-90.
Human Glucagon Prohormone Schematic
Amino Acid Sequence of Glucagon Precursor (Human)
Sequence Comparison between Glucagon, GLP-1 and GLP-2
Bradford et al. Dietary unsaturated fatty acids increase plasma glucagon-like peptide-1 and cholecystokinin and may decrease premeal ghrelin in lactating dairy cows. J Dairy Sci. 2008 Apr;91(4):1443-50.
Lee et al. Glucagon-Like Peptide-1 Gene Therapy in Obese Diabetic Mice Results in Long-Term Cure of Diabetes by Improving Insulin Sensitivity and Reducing Hepatic Gluconeogenesis Diabetes. 2007 Jun;56(6):1671-9.
Liu et al. Prolonged remission of diabetes by regeneration of beta cells in diabetic mice treated with recombinant adenoviral vector expressing glucagon-like peptide-1. Molecular Therapy. 2007; 15: 86-93.
Laferrère et al. Incretin Levels and Effect Are Markedly Enhanced 1 Month After Roux-en-Y Gastric Bypass Surgery in Obese Patients With Type 2 Diabetes Diabetes Care. 2007 Jul;30(7):1709-16.
Granata et al. Obestatin promotes survival of pancreatic beta-cells and human islets and induces expression of genes involved in the regulation of beta-cell mass and function. Diabetes. 2008 Apr;57(4):967-79.
Santoro et al. Enterohormonal changes after digestive adaptation: five-year results of a surgical proposal to treat obesity and associated diseases. Obes Surg. 2008 Jan;18(1):17-26.
Pacheco et al. The effects of duodenal-jejunal exclusion on hormonal regulation of glucose metabolism in Goto-Kakizaki rats. Am J Surg. 2007 Aug;194(2):221-4.
Suzuki et al. Changes in GI hormones and their effect on gastric emptying and transit times after Roux-en-Y gastric bypass in rat model Surgery. 2005 Aug;138(2):283-90.