|
|
|
TRAIL |
Novel TRAIL Isoform and Multimerized TRAIL Mimics for Specific Cell Apoptosis Studies |

|
A proposed model for TRAIL regulating inflammation resolution by modulation of neutrophil lifespan. In WT mice, TRAIL expression by tissue cells, e.g., the respiratory epithelium, can induce apoptosis of recruited neutrophils. In TRAIL-deficient mice, inflammatory cell survival is prolonged until engagement of a constitutive program of neutrophil apoptosis. Exogenous TRAIL accelerates the apoptotic death of neutrophils in TRAIL-deficient and WT mice. |
Figure from : McGrath E.E. et al., J Leukoc Biol. Nov 2011; 90(5): 855–865. doi: 10.1189/jlb.0211062
 |
Virus-host interactions are characterized by the selection of adaptive mechanisms by which to evade pathogenic and defense mechanisms, respectively. In primary T cells infected with HIV, HIV infection up-regulates TNF-related apoptosis inducing ligand (TRAIL) and death-inducing TRAIL receptors, but blockade of TRAIL:TRAIL receptor interaction does not alter HIV-induced cell death. Instead, HIV infection results in a novel splice variant that we call TRAIL-short (TRAIL-s), which antagonizes TRAIL-R2. In HIV patients, plasma TRAIL-s concentration increases with increasing viral load and renders cells resistant to TRAIL-induced death. Knockdown of TRAIL-s abrogates this resistance. We propose that TRAIL-s is a novel adaptive mechanism of apoptosis resistance acquired by HIV-infected cells to avoid their elimination by TRAIL-dependent effector mechanism.
Schnepple DJ, Shepard B, Bren GD et al., J Biol Chem. 2011 Oct 14;286(41):35742-54. doi: 10.1074/jbc.M111.274639. Epub 2011 Aug 22.
|
|
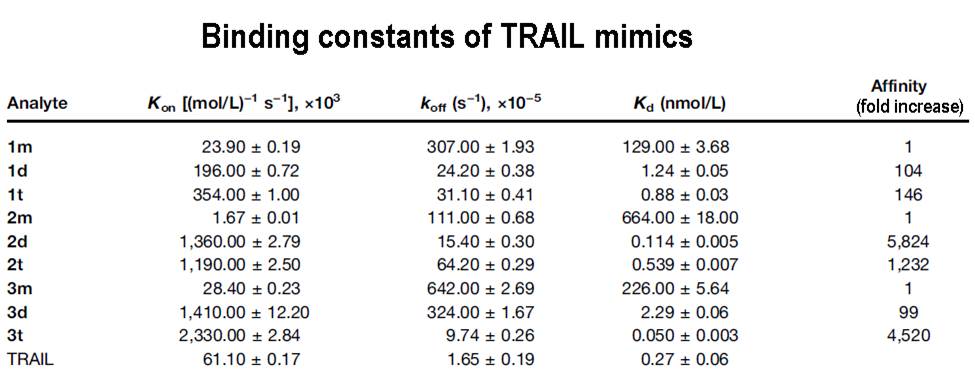
Ongoing clinical trials are exploring anticancer approaches based on signaling by TRAIL, a ligand for the cell death receptors DR4 and DR5. In this study, we report on the selective apoptotic effects of multivalent DR5 binding peptides (TRAIL(mim/DR5)) on cancer cells in vitro and in vivo. Surface plasmon resonance revealed up to several thousand-fold increased affinities of TRAIL(mim/DR5)-receptor complexes on generation of divalent and trivalent molecules, the latter of which was achieved with a conformationally restricted adamantane core. Notably, only multivalent molecules triggered a substantial DR5-dependent apoptotic response in vitro. In tumor models derived from human embryonic kidney cells or primary foreskin fibroblasts, TRAIL(mim/DR5) peptides exerted a cancer cell-selective action that could synergize with resveratrol in a manner independent of p53. In a xenograft model of human colon cancer, a divalent TRAIL(mim/DR5) peptide inhibited tumor growth. Our results offer a proof-of-principle for the development of synthetic small molecules to trigger the TRAIL apoptosis pathway for cancer therapy.
Pavet V, Beyrath J, Pardin C et al., Cancer Res. 2010 Feb 1;70(3):1101-10. doi: 10.1158/0008-5472.CAN-09-2889. Epub 2010 Jan 26.
|
|
|
BACKGROUND:
HIV infected patients have an increased susceptibility to liver disease due to Hepatitis B Virus (HBV), Hepatitis C Virus (HCV), alcoholic, and non-alcoholic steatohepatitis. Clinically, this results in limited options for antiretroviral therapy and accelerated rates of liver disease, causing liver disease to be the second leading cause of death for HIV infected patients. The mechanisms causing this propensity for liver dysfunction during HIV remains unknown.
METHODOLOGY/PRINCIPAL FINDINGS:
We demonstrate that HIV and/or the HIV glycoprotein gp120 ligation of CXCR4 on hepatocytes selectively up-regulates TRAIL R2 expression and confers an acquired sensitivity to TRAIL mediated apoptosis which is mediated by JNK II, but not p38 nor G-proteins.
CONCLUSIONS/SIGNIFICANCE:
These findings suggest that HIV infection renders hepatocytes more susceptible to liver injury during disease states associated with enhanced TRAIL production such as HBV, HCV, or steatohepatitis.
Babu CK, Suwansrinon K, Bren GD et al., PLoS One. 2009;4(2):e4623. doi: 10.1371/journal.pone.0004623. Epub 2009 Feb 27.
|
|
Tumour necrosis factor (TNF) related apoptosis-inducing ligand (TRAIL/APO2L) is a recently identified member of the TNF family, which induces programmed cell death in a variety of neoplastic cell types, but not in most nonneoplastic cells. In this study, we report on the identification of two novel alternative splice variants of TRAIL in neoplastic and non-neoplastic human cells lacking either exon 3 (TRAIL-beta) or exons 2 and 3 (TRAIL-gamma). In both splice variants, loss of exon 3 resulted in a frame shift generating a stop codon with consecutive extensive truncation in the extracellular domain. Ectopic expression revealed a loss of proapoptotic potential for both alternative splice variants. In contrast to the predominantly cytoplasmatic localisation of GFP-tagged TRAIL-alpha and TRAIL-beta, TRAIL-gamma showed an additional association with the cell surface and nuclear membrane. In conclusion, alternative splicing might be involved in fine tuning of TRAIL-induced apoptosis and underlines the complexity of the TRAIL system.
Krieg A, Krieg T, Wenzel M et al., Br J Cancer. 2003 Mar 24;88(6):918-27.
|
|
|
Tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) is a member of the TNF superfamily exerting cytotoxic activities toward tumor cells. Herein, we demonstrate that therapeutic concentrations of interferon alpha (IFNalpha) stimulate the expression of high levels of TRAIL mRNA and the release of elevated amounts of a soluble bioactive form of TRAIL (sTRAIL) in both human neutrophils and monocytes. Supernatants harvested from IFNalpha-treated neutrophils/monocytes elicited, on TRAIL-sensitive leukemic cell lines, proapoptotic activities that were significantly reduced by either a combination of TRAIL-R1/Fc and TRAIL-R2/Fc chimeras or neutralizing anti-TRAIL, anti-TRAIL-R1, and anti-TRAIL-R2 antibodies, suggesting that they were mediated by released sTRAIL acting on both TRAIL receptors. Since diseases such as chronic myeloid leukemia (CML) and melanoma are effectively treated with IFNalpha,we also demonstrate that CML neutrophils and peripheral blood mononuclear cells (PBMCs) cultured with IFNalpha at therapeutic concentrations retain the capacity of releasing sTRAIL, suggesting that CML leukocytes, in vivo, might represent an important source of sTRAIL. In this regard, we show that sTRAIL serum levels as well as leukocyte-associated TRAIL significantly increase in melanoma patients following IFNalpha administration. Collectively, these findings indicate that sTRAIL released by IFNalpha-activated neutrophils and monocytes contributes not only to the immunoregulatory actions but also to the therapeutic activities of IFNalpha.
Tecchio C, Huber V, Scapini P et al., Blood. 2004 May 15;103(10):3837-44. Epub 2004 Jan 15.
|

|
Wajant H, Moosmayer D, Wüest T et al., Oncogene. 2001 Jul 5;20(30):4101-6.
|
|
|
TNF-related apoptosis-inducing ligand (TRAIL) is a typical member of the tumor necrosis factor (TNF) ligand family that is expressed as a type II membrane protein (memTRAIL) and signals apoptosis via the death domain-containing receptors TRAIL-R1 and -2. Soluble recombinant derivatives of TRAIL (sTRAIL) are considered as novel tumors therapeutics because of their selective apoptosis inducing activity in a variety of human tumors but not in normal cells. Using antagonistic antigen-binding fragment (Fab) preparations of TRAIL-R1- and TRAIL-R2-specific antibodies, we demonstrate in this study that TRAIL-R1 becomes activated by both the soluble and the membrane-bound form of the ligand, whereas TRAIL-R2 becomes only activated by memTRAIL or soluble TRAIL secondarily cross-linked by antibodies. Furthermore, we show that the restricted signal capacity of sTRAIL can be readily converted into a fully signal competent memTRAIL-like molecule, i.e. a TRAIL-R2 stimulating ligand, by genetic fusion to an antibody derivative that allows antigen-dependent 'immobilization' of the fusion protein to cell surfaces. We conclude that antibody targeting-dependent activation can be used to design selective therapeutics derived of those ligands of the TNF family that are biologically inactive in their soluble form.
Wajant H, Moosmayer D, Wüest T et al., Oncogene. 2001 Jul 5;20(30):4101-6. |
|
|
|
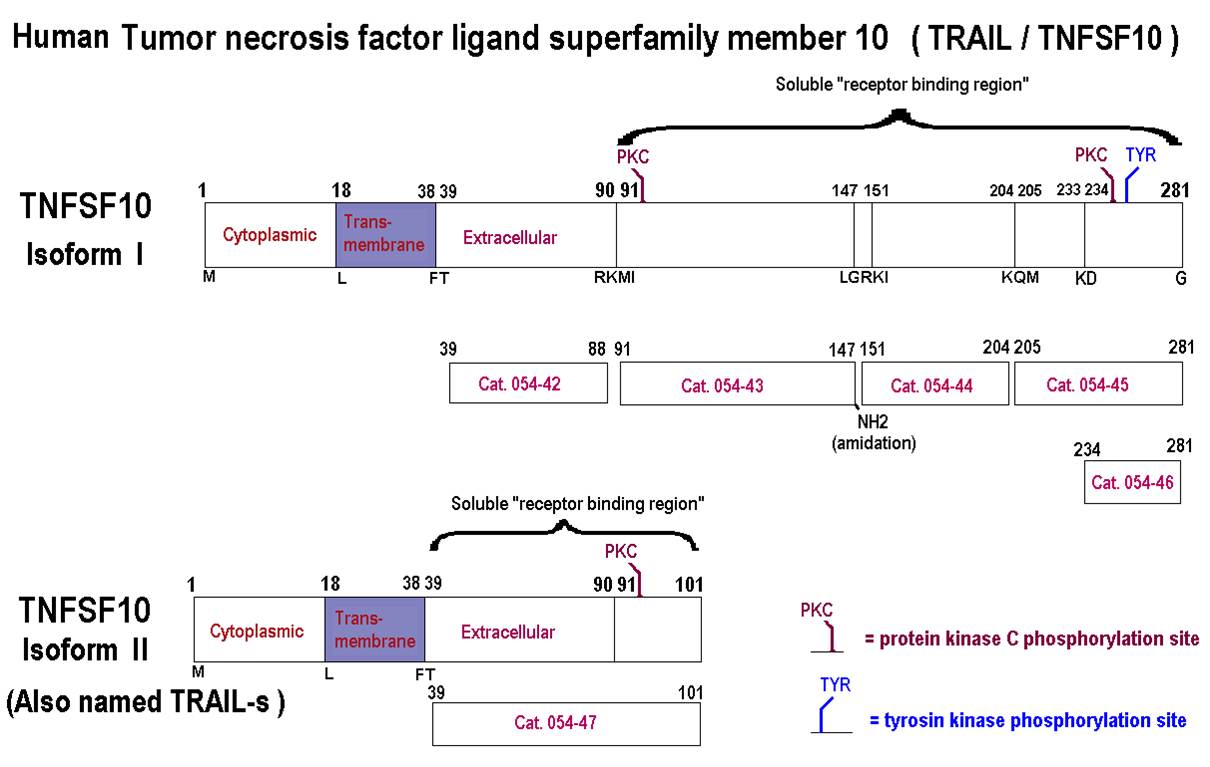
%TRAIL%;%054-47%;%054-6%
|
|
|