Peripheral tissue injury is associated with changes in protein expression in sensory neurons that may contribute to abnormal nociceptive processing. We used cultured dorsal root ganglion (DRG) neurons as a model of axotomized neurons to investigate early changes in protein expression after nerve injury. Comparing protein levels immediately after DRG dissociation and 24 h later by proteomic differential expression analysis, we found a substantial increase in the levels of the neurotrophin-inducible protein VGF (nonacronymic), a putative neuropeptide precursor. In a rodent model of nerve injury, VGF levels were increased within 24 h in both injured and uninjured DRG neurons, and the increase persisted for at least 7 d. VGF was also upregulated 24 h after hindpaw inflammation. To determine whether peptides derived from proteolytic processing of VGF participate in nociceptive signaling, we examined the spinal effects of AQEE-30 and LQEQ-19, potential proteolytic products shown previously to be bioactive. Each peptide evoked dose-dependent thermal hyperalgesia that required activation of the mitogen-activated protein kinase p38. In addition, LQEQ-19 induced p38 phosphorylation in spinal microglia when injected intrathecally and in the BV-2 microglial cell line when applied in vitro. In summary, our results demonstrate rapid upregulation of VGF in sensory neurons after nerve injury and inflammation and activation of microglial p38 by VGF peptides. Therefore, VGF peptides released from sensory neurons may participate in activation of spinal microglia after peripheral tissue injury.
Riedl et al. J Neurosci. 2009 Oct 21;29(42):13377-88.
The effect of five peptides derived from the C-terminal portion of rat pro-VGF (VGF(577-617), VGF(588-617), VGF(599-617), VGF(556-576) and VGF(588-597)) on penile erection was studied after injection into the hypothalamic paraventricular nucleus of male rats. VGF(577-617), VGF(588-617), VGF(599-617) and, to a lower extent, VGF(588-597) (0.1-2 microg) induced penile erection episodes in a dose-dependent manner when injected into the paraventricular nucleus, while VGF(556-576) was ineffective. VGF(588-617)-induced penile erection was reduced by nitro(omega)-L-arginine methylester (L-NAME; 20 microg), by morphine (5 microg) and by muscimol (1 microg), but not by dizocilpine [(+)MK-801; 1 microg], nor by cis-flupenthixol (10 microg) given into the paraventricular nucleus 10 min before the VGF peptide. d(CH2)5Tyr(Me)-Orn8-vasotocin (1 microg) effectively reduced VGF(588-617)-induced penile erection when given into the lateral ventricles but not when injected into the paraventricular nucleus. Immunocytochemistry with antibodies specific for the C-terminal nonapeptide sequence of pro-VGF (VGF(609-617)) revealed numerous neuronal fibres and terminals within the paraventricular nucleus, including its parvocellular components. Here, many immunostained neuronal terminals impinged on parvocellular oxytocinergic neurons. The present results show for the first time that certain pro-VGF C-terminus-derived peptides promote penile erection when injected into the paraventricular nucleus and suggest that, within this nucleus, these or closely related pro-VGF-derived peptides may be released to influence sexual function by activating paraventricular oxytocinergic neurons mediating penile erection.
Succu et al. Eur J Neurosci. 2004 Dec;20(11):3035-40.

A–C, VGF-IR increased dramatically 1 d after SNL (C) and to a lesser extent after sham surgery (B). Corresponding images of YOYO-1 labeling (A'–C') show all cells in the fields of view shown in A–C. D, Quantitative analysis showed a significant increase in the number of VGF-positive neurons 1 and 7 d after SNL in both injured L5 and uninjured L4 DRG. Sham surgery resulted in a more moderate but significant increase in the number of VGF-positive neurons in L5 DRG at day 1 and day 7 and in L4 DRG at day 1. Significant changes are indicated by * when compared with sham and # when compared with naive (*p < 0.001; **p < 0.05; #p < 0.001; ##p < 0.005; ###p < 0.01; one-way ANOVA and Student–Newman–Keuls test for multiple comparisons; n = 4 in each group).
Riedl et al. J Neurosci. 2009 Oct 21;29(42):13377-88.
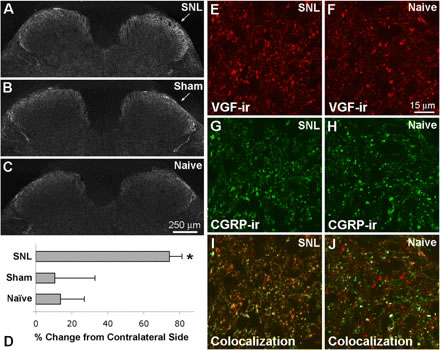
A–C, Three days after SNL, VGF-IR appeared increased within the dorsal horn ipsilateral to SNL (A) compared with sham surgery (B) and naive animals (C). D, Quantitative analysis indicated a significant increase in VGF-IR ipsilateral to SNL compared with sham surgery and naive spinal cord (one-way ANOVA and Student–Newman–Keuls multiple comparisons test, p < 0.05; n = 4 for each group). E–J, Compared with naive spinal cord (F, H, J), VGF-IR (red) was increased after SNL (E, G, I), whereas CGRP-IR (green) appeared unchanged at this time point. The colocalization of VGF- and CGRP-IR in superficial dorsal horn was also increased after SNL as indicated by the presence of more yellow and orange puncta ipsilateral to SNL (I) compared with naive spinal cord (J).
Riedl et al. J Neurosci. 2009 Oct 21;29(42):13377-88.
The inhibitor was injected intrathecally as a 5 min pretreatment before injection of equieffective doses of AQEE-30 (0.3 nmol) or LQEQ-19 (1 nmol). Thermal hyperalgesia was measured 30 and 10 min after intrathecal injection of AQEE-30 and LQEQ-19, respectively. Data are expressed as percentage inhibition of the maximum thermal hyperalgesia (AQEE-30, –2.29 s; LQEQ-19, –2.43 s) detected in a separate control group pretreated with saline.
Riedl et al. J Neurosci. 2009 Oct 21;29(42):13377-88.

Riedl et al. J Neurosci. 2009 Oct 21;29(42):13377-88.