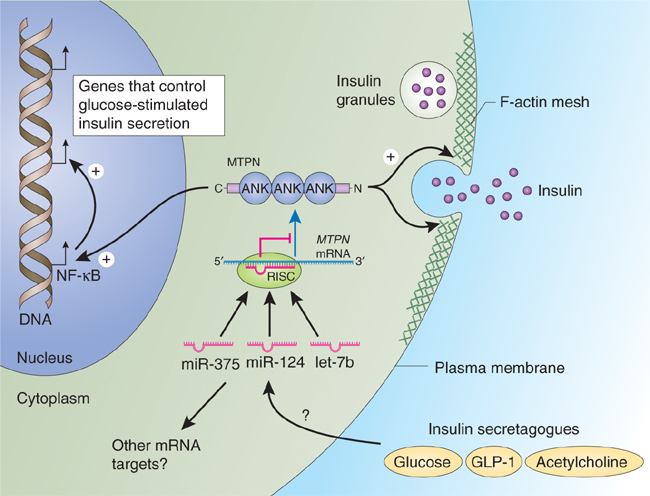
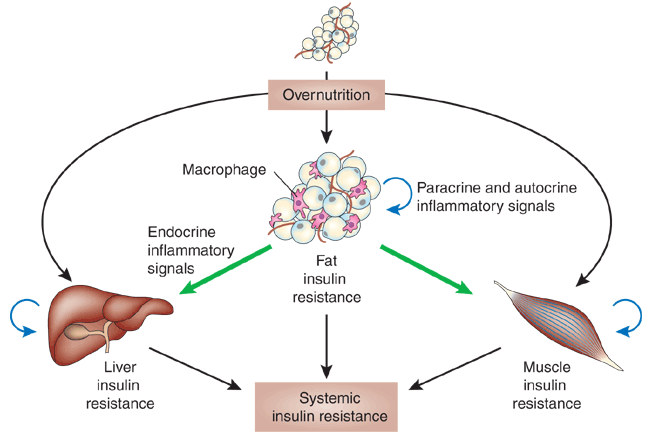
The development of systemic insulin resistance in obesity-induced
inflammation and stress. In obese states, adipose tissue
is under a constant state of metabolic stress, resulting
in the activation of the stress and inflammatory response,
which leads to the accumulation of macrophages. In this
state, adipocytes release cytokines, adipokines and free
fatty acids, which can act in a paracrine or autocrine
fashion to amplify the proinflammatory state within adipose
tissue and cause localized insulin resistance. Adipose
tissue also serves as an endocrine organ whereby these
cytokines, adipokines and free fatty acids travel to liver
and muscle and may decrease insulin sensitivity. In addition
to the adipose tissue–derived factors, stress and
inflammatory signals can arise independently within liver
and muscle, and result in local insulin resistance within
these organs. Carl de Luca
& Jerrold M Olefsky. Nature Medicine 12,
41 - 42 (2006)
The humoral theory of insulin resistance.
In this model, insulin resistance results from pathophysiological
levels of circulating factors that are potentially derived
from several different cell types. The possible role of
adipocytes, macrophages (in adipose tissue, liver and
elsewhere), and hepatocytes is shown, along with secreted
factors that modulate insulin action at the cellular level.
Simon Fenwick. Mitchell A Lazar.
Nature Medicine 12, 43 - 44 (2006)
Insulin regulates glucose and lipid metabolism through
forkhead transcription factors. (a) Under normal
conditions, the insulin pathway regulates Foxo1 and
Foxa2, resulting in regulated glycemia and a balance
of hepatic lipid accumulation and oxidation. (b)
In moderate insulin resistance, reduced activity of
the insulin pathway results in activation of Foxo1,
leading to elevated gluconeogenesis and hyperglycemia.
However, Foxa2 is more sensitive to insulin and therefore
is still repressed, resulting in reduced lipid oxidation
and hepatic steatosis. (c) In severe
insulin resistance, the insulin pathway is barely active,
and the constitutive activation of Foxo1 and Foxa2 result
in elevated gluconeogenesis, hyperglycemia and high
levels of lipid oxidation, leading to ketoacidosis.
Simon Fenwick. Pere Puigserver
& Joseph T Rodgers. Nature Medicine 12,
38 - 39 (2006)
Determination of insulin in a single islet of Langerhans
by high-performance liquid chromatography with fluorescence
detection
Rodents (rat and mouse) have two types of insulin
(insulin I and II; each contains a universal chain A
and a different composition of each type BI chain or
type BII chain). The physiological role for each isomer
is not yet clarified because of the lack of an appropriate
separative determination method for these isomers. Thus,
in this paper, a sensitive and selective HPLC-fluorescence
determination method for the isomers was developed,
which includes derivatization with a fluorogenic reagent
for thiols, 7-fluoro-2,1,3-benzoxadiazole-4-sulfonate,
in the presence of a reducing agent, TCEP, a nonionic
surfactant, n-dodecyl beta-D-maltopyranoside, and EDTA.
The resultant chain A, BI, and BII derivatives were
separated on a reversed-phase column (TSK gel ODS-120T,
250 x 4.6 mm i.d.) with a mobile phase containing 5
mM phosphate buffer (pH 7.0) and were detected at 505
nm with excitation at 380 nm. The detection limits for
chain A, BI, and BII derivatives were 2.2, 3.4, and
3.7 fmol on column, respectively. The method was applicable
to the determination of rodent insulin in a single islet
of Langerhans, and the results indicated its feasibility
for the investigation of the pathophysiological roles
of the isomers in diabetes in the rodent. Toriumi
C, Imai K. Anal Chem.
2002 May 15;74(10):2321-7.
Preproinsulin I and II mRNA expression in adult rat submandibular
glands
Mammalian salivary glands are known to produce a number
of biologically active peptides. The aim of this study
was to extend our previous results showing the presence
of a biologically active insulin-like immunoreactive peptide
in rat salivary glands. In rodents, where two nonallelic
and functional insulin genes are expressed, the co-expression
of both genes seems to be limited to beta-cells of pancreatic
islets or to embryologic developmental processes. We have
investigated the expression of insulin genes in rat submandibular
glands and in a murine immortalized submandibular cell
line, SCA-9. For this purpose, total RNAs were isolated
and submitted to reverse transcription. The cDNAs obtained
were amplified by a nested polymerase chain reaction using
rat preproinsulin I and II primers. Our data show that
both preproinsulin I and II mRNAs are expressed in adult
rat submandibular glands as well as in the SCA-9 cell
line. The identification of salivary gland rat preproinsulin
I and II was confirmed by direct sequencing. These results
provide, for the first time, evidence for the expression
of both preproinsulin I and II mRNA in an extra-pancreatic
tissue from adult rodents.
Egea JC, et al. Eur J Oral Sci.
2000 Aug;108(4):292-6.
Wiegand S., et al. European Journal
of Endocrinology (2004) 151 199–206
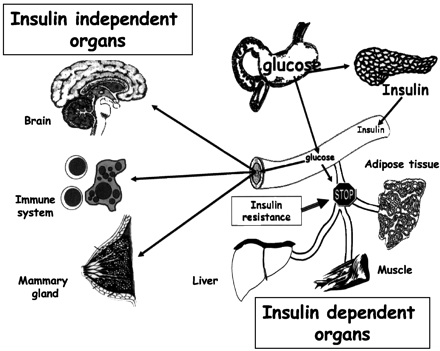
Glucose sensing is among the most conserved pathways in
human evolution, given its vital importance for brain
and immune system metabolism. These two systems are hypothesized
to interact to obtain glucose, no matter the cost, from
adipose, liver, and muscle tissues, even at the expense
of pancreas exhaustion. Evolutively, the price to pay
was very low. Those individuals with cytokine and metabolic
genetic polymorphisms that implicated the best external/internal
defense, preserving glucose for brain, immune system,
placenta, and mammary gland, the classical insulin-independent
tissues, survived and transmitted their genes.
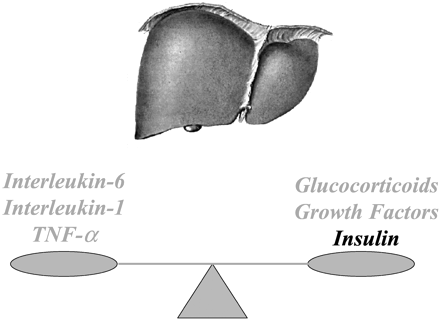
Balance of proinflammatory and antiinflammatory agents
regulating the acute-phase response. An adequate balance
will lead to resolution of the process.