Serum insulin-like growth
factor-I levels and prostatic intraepithelial neoplasia:
a clue to the relationship between Igf-I physiology
and prostate cancer risk
Several lines of evidence indicate
an involvement of brain derived neurotrophic factor
(BDNF) in body weight regulation and activity: heterozygous
Bdnf knockout mice (Bdnf(+/-)) are hyperphagic, obese,
and hyperactive; furthermore, central infusion of BDNF
leads to severe, dose-dependent appetite suppression
and weight loss in rats. We searched for the role of
BDNF variants in obesity, eating disorders, and attention-deficit/hyperactivity
disorder (ADHD). A mutation screen (SSCP and DHPLC)
of the translated region of BDNF in 183 extremely obese
children and adolescents and 187 underweight students
was performed. Additionally, we genotyped two common
polymorphisms (rs6265: p.V66M; c.-46C > T) in 118
patients with anorexia nervosa, 80 patients with bulimia
nervosa, 88 patients with ADHD, and 96 normal weight
controls. Three rare variants (c.5C > T: p.T2I; c.273G
> A; c.*137A > G) and the known polymorphism (p.V66M)
were identified. A role of the I2 allele in the etiology
of obesity cannot be excluded. We found no association
between p.V66M or the additionally genotyped variant
c.-46C > T and obesity, ADHD or eating disorders.
Friedel S, et al. Am J Med Genet
B Neuropsychiatr Genet. 2005 Jan 5;132(1):96-9
Control of hyperphagia prevents obesity in BDNF
heterozygous mice
Reduced levels of BDNF in mice cause obesity and behavioral
abnormalities including increased aggression and hyperactivity.
While it has been shown that the obesity is in part caused
by increased food consumption it is still not clear whether
defects in other mechanisms involved in the control of
body weight homeostasis can also affect this phenotype.
Here we report that mice with reduced levels of BDNF do
not develop obesity and have normal blood glucose levels
if fed over a prolonged period of time the amount of food
that control mice usually consume. Thus, hyperphagia appears
to be the primary cause of obesity development rather
than changes in mechanisms controlling metabolism.
Coppola V, Tessarollo L. Neuroreport. 2004 Dec
3;15(17):2665-8.
Opposite changes in the serum brain-derived
neurotrophic factor in anorexia nervosa and obesity
OBJECTIVE: A role for the brain-derived neurotrophic
factor (BDNF) in the regulation of eating behavior has
been recently demonstrated. Therefore, the possibility
exists that alterations in BDNF production and/or activity
are involved in the pathophysiology of anorexia nervosa
(AN) and obesity. METHODS: We measured morning serum
levels of BDNF in 22 women with AN, 24 women with obesity
(body mass index [BMI] > 30 kg/m2), and 27 nonobese
healthy women. All the subjects were drug-free and underwent
a clinical assessment by means of rating scales measuring
both eating-related psychopathology and depressive symptoms.
RESULTS: As compared with the nonobese healthy controls,
circulating BDNF was significantly reduced in AN patients
and significantly increased in obese subjects. No significant
difference was observed in serum BDNF concentrations
between AN women with or without a comorbid depressive
disorder. Moreover, serum BDNF levels were significantly
and positively correlated with the subjects' body weight
and BMI.CONCLUSION: The BDNF changes observed in AN
and obesity are likely secondary adaptive mechanisms
aimed at counteracting the change in energy balance
that occurs in these syndromes.
Monteleone P, et al. Psychosom Med.
2004 Sep-Oct;66(5):744-8
Reversal of behavioral and metabolic abnormalities,
and insulin resistance syndrome, by dietary restriction
in mice deficient in brain-derived neurotrophic factor
Dietary restriction (DR) extends life span and improves
glucose metabolism in mammals. Recent studies have shown
that DR stimulates the production of brain-derived neurotrophic
factor (BDNF) in brain cells, which may mediate neuroprotective
and neurogenic actions of DR. Other studies have suggested
a role for central BDNF signaling in the regulation of
glucose metabolism and body weight. BDNF heterozygous
knockout (BDNF+/-) mice are obese and exhibit features
of insulin resistance. We now report that an intermittent
fasting DR regimen reverses several abnormal phenotypes
of BDNF(+/-) mice including obesity, hyperphagia, and
increased locomotor activity. DR increases BDNF levels
in the brains of BDNF(+/-) mice to the level of wild-type
mice fed ad libitum. BDNF(+/-) mice exhibit an insulin-resistance
syndrome phenotype characterized by elevated levels of
circulating glucose, insulin, and leptin; DR reduces levels
of each of these three factors. DR normalizes blood glucose
responses in glucose tolerance and insulin tolerance tests
in the BDNF(+/-) mice. These findings suggest that BDNF
is a major regulator of energy metabolism and that beneficial
effects of DR on glucose metabolism are mediated, in part,
by BDNF signaling. Dietary and pharmacological manipulations
of BDNF signaling may prove useful in the prevention and
treatment of obesity and insulin resistance syndrome-related
diseases.
Duan W, et al. Endocrinology. 2003 Jun;144(6):2446-53
Conditional deletion of brain-derived neurotrophic
factor in the postnatal brain leads to obesity and hyperactivity
Brain-derived neurotrophic factor has been associated
previously with the regulation of food intake. To help
elucidate the role of this neurotrophin in weight regulation,
we have generated conditional mutants in which brain-derived
neurotrophic factor has been eliminated from the brain
after birth through the use of the cre-loxP recombination
system. Brain-derived neurotrophic factor conditional
mutants were hyperactive after exposure to stressors and
had higher levels of anxiety when evaluated in the light/dark
exploration test. They also had mature onset obesity characterized
by a dramatic 80-150% increase in body weight, increased
linear growth, and elevated serum levels of leptin, insulin,
glucose, and cholesterol. In addition, the mutants had
an abnormal starvation response and elevated basal levels
of POMC, an anorexigenic factor and the precursor for
alpha-MSH. Our results demonstrate that brain derived
neurotrophic factor has an essential maintenance function
in the regulation of anxiety-related behavior and in food
intake through central mediators in both the basal and
fasted state.
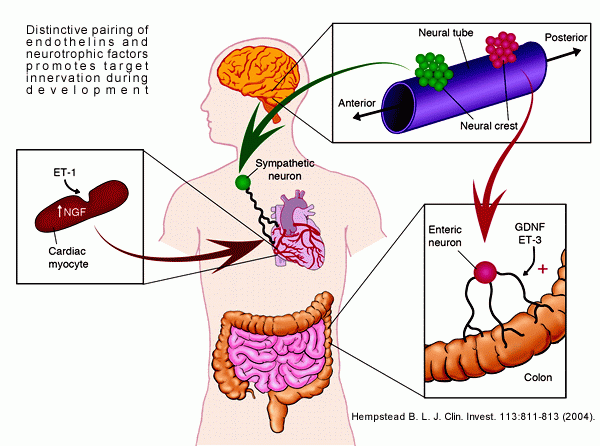
Distinctive pairing of endothelins and neurotrophic
factors promotes target innervation during development.
Although the sequential action of ET-1 to induce myocyte
production of NGF is suggested to promote sympathetic
innervation of the heart, more complex interactions of
ET-3 and GDNF occur in the patterning of the enteric nervous
system.
Barbara L. Hempstead. J. Clin. Invest. 113:811-813 (2004)
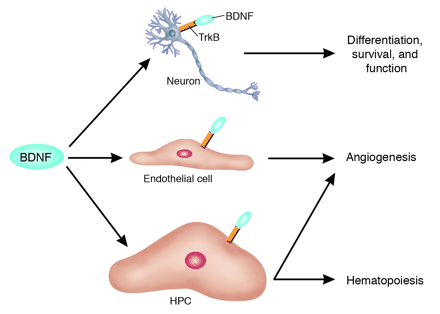
The functional pleiotropy of BDNF extends beyond the
nervous system. BDNF, similar to NT-4, exerts its complex
signaling effects via TrkB that is present on neurons.
These effects include the modulation of neuronal differentiation,
survival, and function. In this issue of the JCI, Kermani
et al. (10) report a more complex role for this neurotrophic
factor: BDNF can mobilize TrkB+ hematopoietic precursor
cells (HPCs) for both hematopoiesis and tissue neovascularization.
In addition, BDNF can promote angiogenesis by directly
interacting with TrkB expressed on ECs. Dan G. Duda and
Rakesh K. Jain. J. Clin. Invest. 115:596–598 (2005)
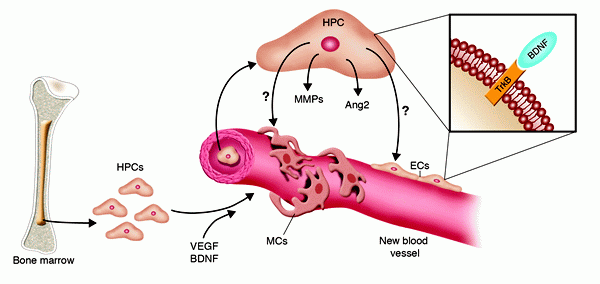
Cellular and molecular mechanisms of BDNF-induced neovascularization.
BDNF was recently implicated in new vessel formation,
in both mouse embryos (8) and adult mice (10). In adults,
the formation of new vessels in response to BDNF overexpression
is the result of both direct effects on TrkB expressed
by tissue-resident ECs and the recruitment of TrkB+VEGFR2+CD11b+Sca1+
myeloid HPCs. The latter cells may indirectly promote
neovascularization by releasing various factors, including
Ang2 and MMPs. Nevertheless, a direct involvement of myeloid
HPCs in vessel formation cannot be excluded, as they also
have the potential to acquire an EC or mural cell (MC)
phenotype. Dan G. Duda and Rakesh K. Jain. J. Clin. Invest.
115:596–598 (2005)
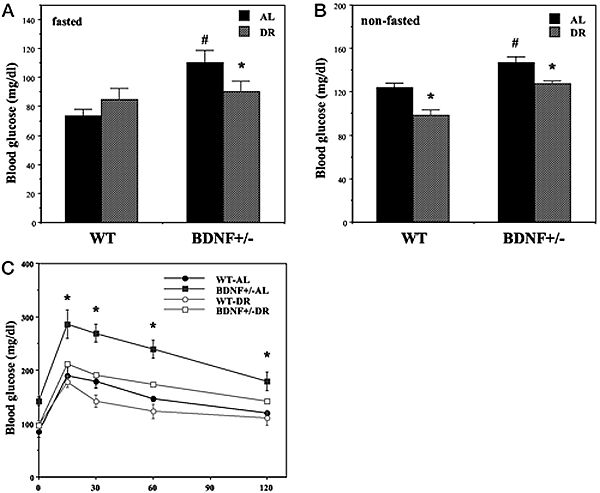
Hyperglycemia and impaired glucose tolerance in BDNF+/-
mice are normalized by DR. Wild-type (WT) and BDNF+/-
mice were maintained for 3 months on AL or DR feeding
regimens. A and B, Glucose concentrations were measured
in blood samples taken after an overnight fast (A) or
during feeding conditions (B). Note that the scales for
the glucose concentrations in the two graphs are different.
*, P < 0.01 compared with the value for the same genotype
of mice fed AL; #, P < 0.05 cmpared to the WT-AL value.
C, Mice were administered an oral bolus of glucose (2
g/kg) and the glucose concentration in blood samples taken
at the indicated times was determined. *, P < 0.01
compared with the value for each of the other three groups
at that time point. Values are the mean and SEM of measurements
made in 8–10 mice per group. Statistical comparisons
were made using ANOVA and Scheffé’s post hoc tests.
Duan W, et al. Endocrinology. 2003 Jun;144(6):2446-53
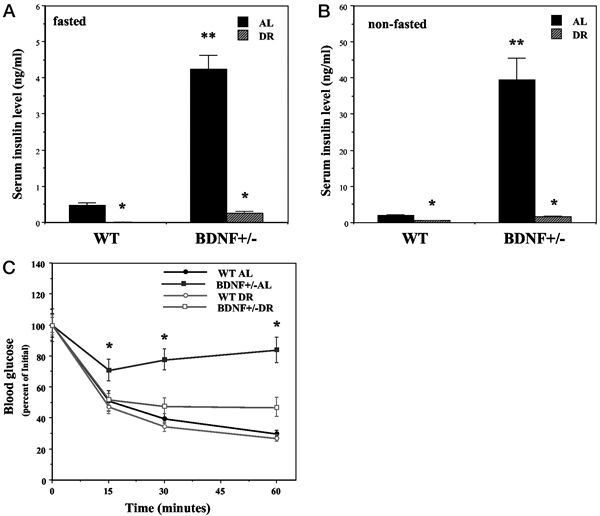
Mice with reduced BDNF levels exhibit insulin insensitivity
that is normalized by DR. Wild-type (WT) and BDNF+/- mice
were maintained for 3 months on AL or DR feeding regimens.
A and B, Insulin concentrations were measured in blood
samples taken after an overnight fast (A) or during feeding
conditions (B). *, P < 0.001 compared with the value
for the same genotype of mice fed AL; **, P < 0.001
compared with the WT-AL value. C, Mice were administered
insulin (1 U/kg) and the glucose concentration in blood
samples taken at the indicated times was determined. *,
P < 0.01 compared with the value for each of the other
three groups at that time point. Values are the mean and
SEM of measurements made in 8–10 mice per group.
Statistical comparisons were made using ANOVA and Scheffé’s
post hoc tests. Duan W, et al. Endocrinology. 2003 Jun;144(6):2446-53
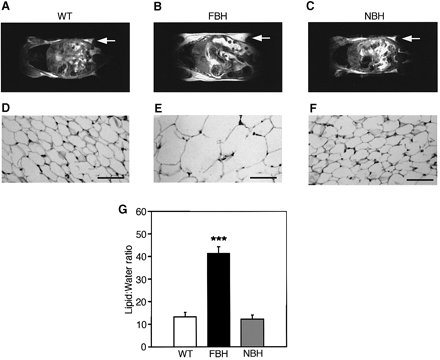
Fat BDNF heterozygous mutant (FBH) mice are obese and
show adipocyte hypertrophy. NMR images (A-C) of whole-body
horizontal sections are taken at the approximate midline.
White density denotes fat, and arrows point to areas of
significant fat accumulation in retroperitoneal fat pads.
(D-F) Fat histology of inguinal fat pads with the bar
representing 70 µm. Quantification of lipid is based on
spectroscopy of lipid and water peaks (G) (see Materials
and methods). Values represent the mean ± SEM. Wild-type
(n = 15), FBH (n = 11) and NBH (n = 8) are labeled accordingly.
Steven G. Kernie, Daniel J. Lieb and Luis F. Parada. The
EMBO Journal Vol. 19, pp. 1290-1300, 2000
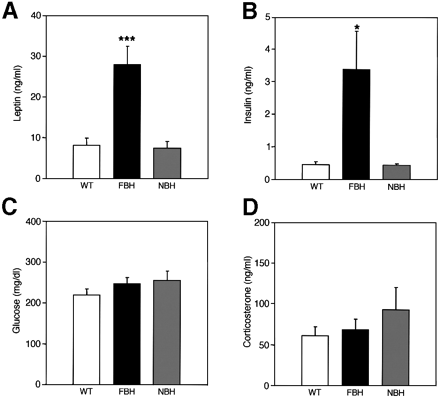
FBH mice are leptin and insulin resistant. Plasma concentrations
of leptin (A), insulin (B), glucose (C) and corticosterone
(D) were measured from wild-type, FBH and NBH mice. Blood
was drawn and processed as described in Materials and
methods. Each value represents the mean ± SEM of 9-23
(wild-type), 12-24 (FBH) and six (NBH) mice. ***p <0.001;
*p <0.05. Steven G. Kernie, Daniel J. Lieb and Luis
F. Parada. The EMBO Journal Vol. 19, pp. 1290-1300, 2000
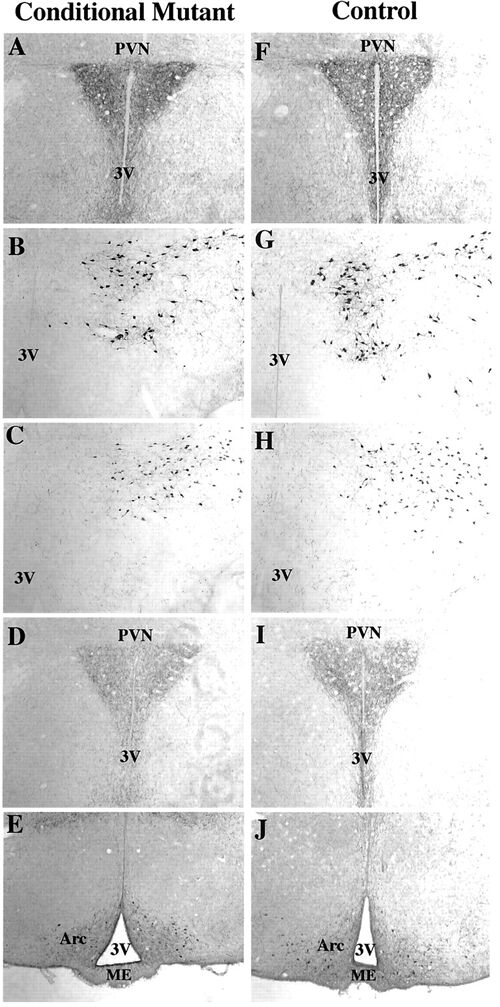
Immunohistochemical Analysis of the BDNF Conditional
Mutant Hypothalamus
Immunohistochemical delineation of NPY (A and F), MCH
(B and G), orexin (C and H), AGRP (D and I) and alpha-MSH
(E and J) in the hypothalamus of conditional mutant
(A–E) and control (F–J) mice. No differences
could be distinguished between the animal groups (n
= 3). PVN, Paraventricular nucleus; 3V, third ventricle;
Arc, arcuate nucleus; ME, median eminence. Rios M.,
et al. Molecular Endocrinology 15 (10): 1748-1757